黄志仁,大麦育种专家。发现大麦互为连锁的矮秆(dw-x) 和多节 (m4) 两个隐性基因并定位于第4染色体上,对大麦新矮源的发现和利用进行论证。育成多个具有良好的利用价值和推广前景的大麦新品种 “扬饲麦1号”、“扬饲麦3号” 和 “扬农啤2号”。为我国大麦育种研究做出了贡献。
黄志仁,祖籍江苏省南通市启东县,1934年2月11日出生于江苏省盐城市大丰县利民村农民家庭。甲午海战中国惨败后,南通张謇实业救国的第一步就是兴办纱厂。为此利用苏北盐场荒凉萧索的机遇,大兴废灶植棉的浪潮,大批启海农民涌入苏北盐垦新棉区,黄志仁就是其中第二代移民。其父母恪守再苦也要让男儿多读书的规教,黄志仁及其兄弟接受了普及教育。抗日战争胜利后,1946年初,他在新四军兴办的台北中学接受为期半年的初中教育,至当年初秋旋因内战爆发而辍学,嗣后曾在米店学徒。
中华人民共和国成立后,黄志仁于1949年11月在苏北行署农水处测量队参加工作,其时年方16岁,后经治淮工程水利训练班培训,在苏北灌溉总渠江都县邵伯工区担任工程员。1951—1957年他奔波于苏北和江苏省治淮指挥部的水利工地,为兴水赈农而努力工作,曾被嘉奖为苏北灌溉总渠工程三等功臣(1952)、江苏省治淮指挥部三河闸工程三等模范 (1953)。其后他被调任宣教干事、电影放映队队长和宣教科科员。他于1955年加入中国共产党。
20世纪50年代的大规模经济建设时期,黄志仁求知若渴,在党和政府 “向科学进军” 口号的感召下,利用业余时间补习初高中主要课程,并立志走上了自学迎考的艰苦历程。他于1957年以同等学力考入原苏北农学院农学专业就读。1960—1961年曾任系团总支书记。1961年大学本科毕业拟留校任职,但他渴望深造,经考试被录取为北京农业大学研究生,在刘中宣先生悉心指导下,攻读作物育种专业。1964年他学成返回苏北农学院农学系遗传育种教研室任教,先后任教研室副主任和教师党支部书记,1979年任讲师,1986年任副教授,1980年兼任江苏农学院大麦研究室副主任,1987年任遗传育种教研室主任,1990年任江苏农学院和扬州大学农学院教授。学习和从教期间他先后荣获优秀青年、学生、优秀团员、党员和先进个人等称号。自1996年起享受国务院政府特殊津贴。
几十年来,黄志仁心无旁骛,积微成着,兢兢业业,一直从事作物遗传育种的教学和研究工作。他多次参与 “七五”、“八五” 和 “九五” 国家农业科技攻关大麦育种项目的起草与论证,并承担国家科委和江苏省科委多项大麦应用基础理论研究课题,其中1991年主持国家“八五”科技攻关项目的大麦杂种优势利用专题,为组织全国大麦雄性不育 “三系” 研究做出了有益贡献。1993年以来他3次荣获江苏省科技奖和1次农业科技奖,3次荣获扬州市科技奖和2次论文奖以及学院多项科技奖。他共发表论文70余篇,参加主编和编审 《中国大麦文集》 第1—5集,主要论着有 《中国大麦学》 (副主编,中国农业出版社出版,1996)、《中国种子市场学》 (第二作者,中国农业大学出版社出版,1998),参加编写《作物育种基础知识》 (江苏科技出版社出版,1981)、《中国大麦品种志》 (中国农业出版社出版,1989)、《江苏麦作科学》 (江苏科技出版社出版,1994)、《21世纪中国农业科技展望》 (山东科技出版社出版,1993) 等共10余部。参加编写《中国大百科全书》 和 《中国农业百科全书》等有关条目。1995年由他主持的作物育种学课程荣获江苏省优秀课程二等奖。经他倡议和努力,于20世纪末开设作物种子专业,在校内首次开设种子学课程,刻意培养和策励青年教师上岗执教。
他参加中国作物学会大麦专业委员会筹建工作,积极组织全国第一次大麦学术讨论会于1985年在扬州召开。自1986年迄今,任大麦专业委员会第1、2届副主任委员和第3、4届常务副主任委员,负责南方,尤其是江苏、浙江、上海地区的学会活动。任中国作物学会第5届理事会理事。出席第5届 (日本,1986)、第6届 (瑞典,1991) 和第8届 (澳大利亚,2000) 国际大麦遗传学论文报告会。他倡导学会应该广泛团结同仁,积极组织和发展学术活动,他认为大麦学术活动的振兴,应与大麦为原料的啤酒酿造、饲料加工和食品产业的大发展同呼吸,因此他一直主张大麦研究始终应多向发展,力促大麦品种改良应及时多向延伸,并倡议大麦研究要开拓思路,积极为大麦产业化服务。他主持南方饲用大麦品种区域试验,对我国饲料大麦的发展做出了贡献。
麦类产量性状遗传规律和产量早代测试及育种应用
黄志仁在攻读研究生期间,开展小麦品种间杂交后代(F
1—F
5) 产量性状遗传及早代产量测试研究,即能否通过杂种早代测验取舍组合,从而提高育种效率。由于受 “文化大革命”影响,黄志仁的研究结果未能及时发表,但有关杂种组合早代测验的初步结论曾被张树榛编入蔡旭主编的 《植物遗传育种学》专着中 (科学出版社出版,1988,第2版,第13章)。
1968年,黄志仁 “业余” 恢复大小麦育种研究。经过数年辛劳,他从特早熟皮大麦推广品种2—14中选育出更早熟1—2天、矮秆、抗病、抗倒伏力强而高产的裸大麦 “8023白壳” 和“8023紫壳”。其中 “8023白壳” 子粒蛋白质含量16.20%、赖氨酸含量0.47%,为优质食用品种,该品种曾荣获农学院科技进步一等奖。在20世纪80年代前后,该两品种在棉麦套作和稻麦三熟制中发挥过良好作用。
1980年江苏农学院成立大麦研究室,黄志仁作为主要负责人将工作重点转向大麦遗传育种研究。他认为性状遗传和种质资源研究是改进育种方法和提高育种效率的重要支柱。他把小麦杂种早代的测试结论延伸下去,通过大麦产量性状和产量比较研究,回答早代组合产量比较能否有效地选留高产组合,在高产组合中能否比中、低产组合产生出较多的高产基因型等问题。
他针对有关产量的12个性状做大麦双列杂交试验,开展遗传力、配合力和亲子相关研究,根据上述性状均以加性效应为主的特点,他提出按h
N2、V
gc和GCV高低将性状分类以进行针对性选择,从而提高了组合确定的优选效果。他每年以2个世代的速度,共历经5年对F
2-F
6共9个世代的杂种组合及其亲本进行产量比较试验,再从产量高、中、低的各1个组合中随机抽取30个单株对其F
7-F
9做株系产量比较试验。试验结果表明,尽管年度间的产量水平差异很大,但各年内组合间的产量高低顺序基本一致,即使F
2和F
3仅经历1—2年产量比较,对组合产量若以
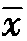
+1/2LSD作为选择标准, 仍可有效地选出高产组合。 遗传分析表明,杂交组合的产量主要受制于亲本的加性作用,而受父母本的连应作用影响较小。组合产量的高低与穗粒重和穗粒数相关密切,因而可把此两性状视为选择高产组合的辅助指标,也有利于田间目测选择。黄志仁采取此法曾选育出684等优良品系。从90个株系的3年产量比较试验中发现,高产组合的30个株系的平均产量和最高株系产量及最低株系产量均高于中、低产组合,因而证明通过早代产量比较测验能有效地选出高产组合,而高产组合比中、低产组合含有较多的高产基因型。黄志仁在小麦领域的先行探索,继而在大麦领域长期系统的研究表明通过相关性状鉴定,优先确定高产组合,以充分表达加性遗传效应,达到加速高产育种目的的上述结论,被同行认为在理论表述上尚属首次报道。
在自花授粉作物杂交育种程序中,是采用传统的系谱选择法,还是采用集团混合选择法,一直是颇有争论的课题。目前至少在大麦育种中尚未采用混合选育法。他们的工作证实,杂交育种的早世代就应倾心选留穗粒数、穗粒重并兼顾选择千粒重方面的优良类型,同时适时淘汰如生育期、株高和抗性等明显不良材料,如此历经3—4个世代混合种植,就可取得既有一定数量又具相对遗传同质化的种质群体。黄志仁认为这就是以高产改良为目标,通过集团混合选择而产生的高产优良基因型。这些结论曾被第5届国际大麦遗传学论文报告会 (日本,冈山,1986) 选录,入编Barley Genetics (1987,V : 1015—1023),在国内收录于《中国大麦文集》 (第1集,1986)。
20世纪80年代后期,针对我国啤酒大麦育种状况,黄志仁搜集江、浙、沪品种,配置双列杂交试验,经遗传分析指出,当时应用的新品种 (系) 期望优势偏低的重要原因,与遗传距离偏小即与亲本来源匮乏有关。他提出必须扩大种质资源的引进力度,在组合优选上多下工夫。在产量构成因素中,二棱大麦应该调整选择重点,更多注意单株穗数和每穗粒数对产量的改良作用,并对当时主栽品种作为亲本利用应该如何发挥其遗传特点提出可臻参考的意见。
不同棱型间大麦杂交应为多向育种提供更为广阔的选择背景和改良途径,该领域工作一直引起黄志仁的注意。他们的观察指出,跨棱型杂交时,F
2代性状分离明显偏大,遗传变异潜力和期望选择效果也更具诱人前景,为此必须相应放大F
2代种植群体,以表现可观的分离趋向和选择效果。通过不同棱型间杂交,对于降低株高、增加穗长、提高单株穗粒数,均比同棱型品种间杂交具有较好的改良效果。
大麦子粒品质性状遗传变异研究
谷粒蛋白质和赖氨酸含量是大麦育种中必须区别对待的理论问题。啤酒酿制大麦在保持子粒粉质化的同时,蛋白质含量亦宜控制在适量范围内而不能偏高; 而人类食用或家畜饲用加工大麦则要求高蛋白质含量,尤其必须的赖氨酸含量偏高。鉴于以往上述遗传变异分析方法的不完善和不确定,因而判定手段已成为制约子粒品质改良的障碍。由莫惠栋教授建立的胚乳3N遗传分析模型有利于澄清以往子粒品质遗传研究中某些概念不清或视听模糊的观念。
黄志仁以大麦谷粒品质性状为对象,最早引用上述模型并发表研究成果指出,大麦谷粒蛋白质含量遗传原则上符合加性一显性模型,且加性效应更为普遍。模型分析认定,着名双高品种Hiproly的高蛋白性状受隐性基因支配,而低蛋白性状为显性遗传。由此提出当以子粒高蛋白 (如饲用或食用) 为选择目标时,在早世代应尽量保留隐性基因在待选群体中的存在概率,以免隐性高蛋白基因过早被淘汰。在这种思想指导下,他们育成 “扬饲麦3号” (泾大1号/Hiproly),不仅子粒蛋白质含量高达15.56%,而且高度抗病,产量高达6 750千克/公顷,成为优质饲用大麦,经审定并在江苏、湖北大面积推广。在啤酒大麦选育时,由于选择目标为相应控制蛋白质含量,因此早代就必须优选低蛋白质材料 (显性),以提高选择效果。研究成果曾在第6届国际大麦遗传学讨论会上 (瑞典,赫尔辛堡,1991) 交流,并入编Barley Genetics (1991,Ⅵ: 452—454)。
黄志仁曾对大麦谷粒蛋白质性状在亲子代 (F
1—F
3) 间的杂种优势及其配合力做过考察,证明亲子代间的一般配合力方差是特殊配合力方差的3—4倍 (2.9—4.5),且一般配合力与亲本的表型值相关度极高 (r=0.92—0.97)。经遗传分析表明,一般配合力虽含有杂种优势部分,但仍以品种效应为主,故亲本表型值与一般配合力的相关达极显着水平,他提出通过亲本的表型值即可预测其一般配合力,并主张以此作为确定亲本的指标; 而特殊配合力虽与杂种优势相关较密切 (r=0.93—0.95),但其变异远小于一般配合力,且不同年份或世代间的相关度亦不甚稳定,故作为亲本组合评价的意义是有限的。此外由于杂种优势很容易测定,而特殊配合力尤须设计专门试验,故不如直接测定杂种优势更为方便。
高蛋白和高赖氨酸并存即 “双高” 选育,已成为饲用和人类食用品种选育的期望性选择目标。问题的复杂性恰在于,在谷粒库容条件下的 “双高” 选择指标往往相互抵牾,形态判断的诸多矛盾亦有待澄清。1997—2001年间他们的研究表明,大麦谷粒的蛋白组成为盐溶蛋白 (占30.96%)、醇溶蛋白(36.76%) 和谷蛋白 (32.28%)。其中盐溶蛋白所占蛋白比例虽略低,但赖氨酸却占谷粒总赖氨酸的61.11%,黄志仁提出在“双高” 改良中,于顾及高蛋白总量选择的同时,必须叠加高盐溶蛋白的选择比例,可望达到精确实现 “双高” 改良的目的。国外有学者在高赖氨酸育种中曾提出选择大胚的方法,他们在皮大麦中发现,谷粒赖氨酸含量与谷粒胚比 (胚占谷粒总干重的比值) 呈显着正相关,而在裸大麦中这种相关不存在。但胚比与千粒重在皮、裸大麦中均呈负相关。这就是说,在皮大麦育种中,胚比可以作为精选高赖氨酸材料的形态辅助选择指标,但改良材料的粒重可能偏低。而这种指标在裸大麦选育中却未必适用。《麦类作物学报》第20卷(2000) 和第22卷(2002) 对此曾有系列报道。
矮秆多节资源的遗传学研究
20世纪80年代初,黄志仁从上海市农业科学院引入二棱大麦早熟3号的矮秆突变体76-2104,该材料株高50厘米以下,主茎8—12节,缩节而丛生,千粒重仅28克,可育率70%左右。以往对大麦矮秆和多 (缩) 节性状,认为是1对隐性基因的多效性 (即一因多效) 所致。黄志仁采用常规品种与76-2104做多次正、反交,再对F
2代矮秆和多节性状的分离趋势试做似然函数 (x
2) 测验,发现在杂交后代中出现比例很低的多节高秆和正常节矮秆的非亲本重组类型,从而认定多节和矮秆并非一因多效,而是分别由互为连锁的两对隐性基因所控制。此后黄志仁从瑞典Lundqvist U教授处引入在7对染色体上各有2个标志性状,共组成14个标记基因系的大麦材料为父本,分别与76-2104杂交,对其性状分离值做最大似然法分析,发现与以往大麦矮秆基因被定位于第2或第3染色体 (m2、m3) 迥然不同的是,决定76-2104矮秆和多节性状的两对基因,均位于第4染色体上,为全新发现。根据命名惯例,黄志仁等将矮源76-2104的多节基因定名为m4,而将矮秆基因定名为dw-x。
由于76-2104的矮秆与多节性状并非 “一因多效”,而是由两对隐性基因所决定,尽管它们位于同一染色体上紧密连锁,但通过杂交可能分离已成事实,因此76-2104作为品种改良的新矮源,其利用前景是可观的。人们可以保留利用其矮生性,或者通过杂交改良而重塑半矮生,同时,通过杂交途径也可以剔除其多(缩) 节丛生及部分不育等不良性状。有关新矮源的报道提交第6届国际大麦遗传学会议 (瑞典,赫尔辛堡,1991) 并被编入Barley Genetics (1991,Ⅵ: 308—309)。中文报道发表于《作物学报》 [1998,24 (6):768—774; 1999,25 (2): 150—156]。
潜心种质资源研究 育成大麦新品种
黄志仁认为只有开拓资源研究,才能开辟亲本利用的新途径。他主张现代大麦育种更要从地域的多样性和用途的多向性出发,因地域和用途 (加工) 的差异,而有意识地开展多向选育,以期不断推出比现有品种有所改良的新类型。
黄志仁等曾对828份 (其中国内材料540份,国外288份)啤用大麦的16项啤用加工品质和相关农艺性状做数年考察和测定,编制出我国第一部啤用大麦品质性状种质资源目录。发现在我国啤用大麦主产区内,不仅在育成品种中存在优良种质资源,而且在传统的地方品种中,亦有低蛋白和高糖化力的优良啤用种质资源。在引进种质中,来自日本的资源无论在品质性状或适应性方面,均比欧美品种更具可利用价值。
为开展大麦黄花叶病抗性资源调研,曾历时3年借助一块发病极重的试验田,对1500余份国内外品种资源进行筛选。考虑到病毒株系的地区差异,又将初步鉴定的抗病材料分别种植于扬州、吴县和如东等三地做多点试验,最后鉴定出314份抗源(抗性为0—1级),其中地方品种145份,改良品种61份,引进的和外国品种100份; 若以类型划分则二棱皮麦64份,二棱裸麦10份,多棱皮麦157份,多棱裸麦83份。调查表明江苏地方品种存在着较多的抗源,为抗病新品种的选育提供了可臻利用的亲本。黄志仁通过多代双列杂交试验进行黄花叶病抗性遗传学研究,发现在不同品种中抗性因子既受1—2对主效基因支配,也受多对微效基因所控制,其抗性的显隐性规律因组合而异,抗性遗传的表现较为复杂。从育种角度出发,他认为在发病区对新品种抗性改良的要求应恰如其分,只要农艺性状好,发病轻而又不影响产量的材料也可利用。他提出大麦抗黄花叶病的育种程序应为:F
1代种植于重病区,以检验其抗性的显隐性和发病程度;F
2代宜种植于无病区,以不受病害影响而优选丰产性和综合性状良好的基因型;F
3代再种植于重病区,为抗性选择奠定基础;F
4代种植于无病区,至分离稳定后再进入病圃做最后抗性鉴定。按照如上程序他们育成的新品种对大麦黄花叶病均属高抗。
黄志仁科研群体向社会共提供6个大麦新品种,其用途多异,在不同地域空间发挥了积极作用。除上述裸大麦 “8023白壳” 和 “8023紫壳”,皮大麦 “扬饲麦3号” 外,还育成 “扬饲麦1号” (西引2号/菲特210),于1998年审定,比西引2号增产10%,蛋白质含量13.7%,取代西引2号而成为饲用大麦的主栽品种; 啤用大麦苏农21选自鬼怒1号,于1993年审定,蛋白质含量11.08%,浸出率80.14%,为江苏省第一批优质啤麦育成品种; 另一啤用品种为扬农啤2号 (Q318/S191/苏引麦2号),于2000年审定,株高仅70—75厘米,抗倒伏力强,6 000千克/公顷以上,高抗大麦黄花叶病,兼抗条纹叶枯病,浸出率79%,千粒重40克以上。该品种以其抗倒、高产、优质、抗病等综合性状优良而在啤用品种中表现更为突出。审定后3年内已迅速推广达9万公顷。他所育成的饲麦和啤麦品种在江苏、河南、湖北等地推广种植,累计面积已达54万公顷以上,为我国大麦扩大种植和作物结构调整做出了应有的贡献。
简历
1934年2月11日 出生于江苏省大丰县利民村。
1946—1949年 在大丰县台北中学学习 (半年) 和米店学徒。
1949年11月—1953年8月 在苏北行署农水处和苏北治淮指挥部工作。
1953年9月—1957年9月 在江苏省治淮指挥部工作。
1957—1961年 苏北农学院农学专业学习。
1961—1964年 北京农业大学作物遗传育种专业攻读研究生。
1964—1972年 任苏北农学院作物遗传育种教研室助教。
1972—1979年 任江苏农学院作物遗传育种教研室副主任。
1979—1986年 任江苏农学院农学系讲师、大麦研究室副主任 (1980)。
1986—1990年 任江苏农学院副教授、遗传育种教研室主任 (1987)。
1990年至今 任江苏农学院和扬州大学 (合并后更名) 教授。
主要论着
1 黄志仁,龚荐,封福如. 用CO
2示踪研究小麦叶片早衰对籽粒产量的影响. 作物学报,1981,7 (1): 27—35
2 黄志仁. 小麦数量性状的初步研究. 遗传学报,1982,9 (1): 63—70
3 黄志仁. 裸大麦杂种F
2数量性状遗传研究. 中国农业科学出版社,1982,15 (6): 48—49
4 黄志仁. 六棱大麦杂种优势与配合力的研究. 作物学报,1984,10(2): 123—132
5 黄志仁,周学美. 大麦杂种F
2组合产比及其性状遗传. 江苏农业学报,1986,2 (1): 41—43
6 黄志仁,周学美. 二棱大麦杂种F
2产量性状的遗传研究. 中国大麦文集 (一),1986: 153—159
7 黄志仁,周学美. 六棱大麦杂种组合产量比较及产量性状的遗传研究.中国大麦文集 (一),1986: 173—182
8 Huang Z R,Zhou M X. Test of the Yield Potential of Balk Hybrids of Sixrowed Barley and the Inheritance of Yield Character. Barley Genetics,Okayama,Japan. 1987,V: 1015—1023
9 黄志仁,周美学,黄友圣等. 大麦蛋白质含量的遗传研究. 作物学报,1991,17 (1):47—53
10 黄志仁,周美学,黄友圣. 大麦籽粒蛋白质含量的配合力分析. 遗传学报,1991,18 (3): 263—270
11 Huang Z R,Zhou M X,Huang Y S. Genetics studies of grain protein content in Barley. Barley Genetics,Helsingborg Swenden,1991,Ⅵ (2):452—454
12 Huang Z R,Zhou M X,Huang Y S. The inheritance of many-nodes and dwarfism. Barley Genetics,Helsingborg Swenden,1991,Ⅵ:308—309
13 黄志仁,黄友圣,周美学等. 大麦黄花叶病抗性的配合力分析. 中国大麦文集 (三),1993: 73—76
14 黄志仁,高文翔. 大麦主要营养品质性状的遗传研究. 中国大麦文集(三),1993: 81—89
15 黄志仁. 大麦的研究与综合开发. 见: 卢良恕主编.21世纪中国农业科技展望. 济南: 山东科学技术出版社,1993: 405—411
16 黄志仁. 大麦的品种改良. 见: 郭绍铮等主编. 江苏麦作科学. 南京:江苏科学技术出版社,1994: 94—120
17 周美学,黄志仁,许如根. 大麦杂优利用中强优势组合的选择和恢复系恢复能力的研究. 江苏农学院学报,1995 (1): 3—4
18 黄志仁. 第六届国际大麦遗传学会议概况. 见: 周恒主编. 农业科技新发展. 中国农业科学技术出版社,1996: 130—134
19 邵启全,黄志仁等. 大麦遗传. 见: 卢良恕主编. 中国大麦学. 北京:中国农业出版社,1996
20 金银根,黄志仁,周美学等. 不育系与可育系大麦开花特性与小穗结构比较. 作物学报,1996,22 (5): 617—621
21 段发平,黄志仁,黄友圣. 大麦突变体弯秆矮生性状的遗传研究. 作物学报,1997,24 (2): 169—175
22 郭杰,黄志仁,徐大勇等编着. 中国种子市场学. 北京: 中国农业大学出版社,1998
23 黄志仁,周美学,黄友圣等. 大麦杂种组合产量潜力及其选择效果的研究. 见: 王连铮,戴景瑞主编. 全国作物育种学术讨论会论文集.北京: 中国农业科学技术出版社,1998: 201—208
24 黄志仁,段发平,许如根等. 大麦多节矮秆性状的研究: Ⅰ. 大麦多节矮秆性状的遗传特点. 作物学报,1998,24 (6): 765—774
25 徐大勇,黄志仁,黄友圣等. 大麦多节矮秆性状的研究: Ⅱ. 大麦多节矮秆性状的染色体定位. 作物学报,1999,25 (2): 150—156
26 黄友圣,徐大勇,黄志仁等. 大麦黄花叶病抗性的遗传研究. 中国大麦文集 (四),1999: 89—98
27 张恒敢,黄志仁,许如根. 大麦雄性不育三系线粒体DNA的酶切分析. 中国大麦文集 (四),1999: 118—123
28 Huang Z R,Xu R G,Zhou M X et al. Studies on barley cytoplasmic male sterility. Barley Genetics,Adelaide Australia,2000,Ⅷ (2): 19—21
29 黄志仁,许如根,吕超等. 大麦高赖氨酸的遗传与选择研究: Ⅰ. 大麦品种蛋白质组分及赖氨酸含量的分析. 麦类作物学报,2000,26(4): 22—28
30 黄志仁.20世纪大麦生产与科研的回顾. 中国大麦文集 (五),2001:1—7
31 黄志仁,吕超,许如根. 大麦高赖氨酸的遗传与选择研究: Ⅱ. 大麦的赖氨酸含量与胚比及千粒量的相关. 麦类作物学报,2002,22 (4):19—22
32 黄志仁. 大麦育种. 见: 盖钧镒主编. 作物育种学各论. 北京: 中国农业出版社,2005
33 黄志仁. 大麦育种. 见: 盖钧镒主编. 作物育种学各论 (第二版).北京: 中国农业出版社,2006: 107—127